PDF Publication Title:
Text from PDF Page: 007
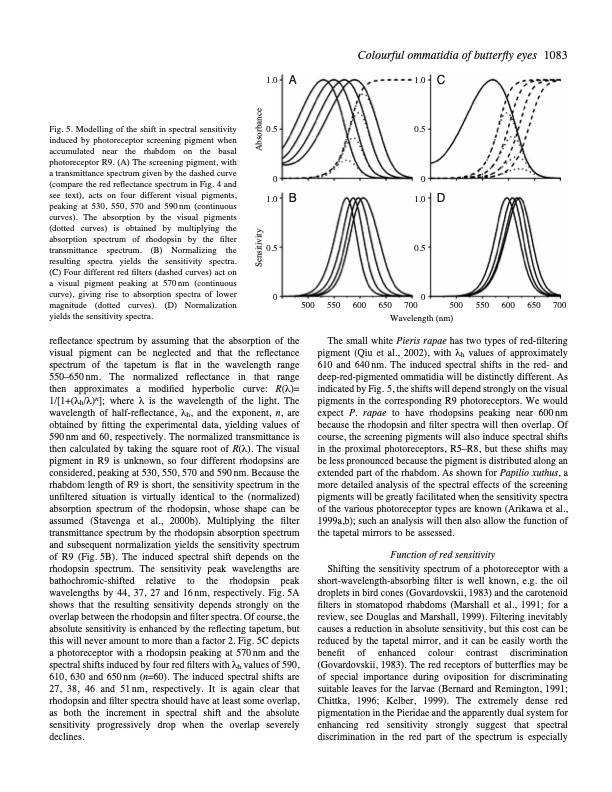
Colourfulommatidiaofbutterflyeyes 1083 Fig. 5. Modelling of the shift in spectral sensitivity induced by photoreceptor screening pigment when accumulated near the rhabdom on the basal photoreceptor R9. (A) The screening pigment, with a transmittance spectrum given by the dashed curve (compare the red reflectance spectrum in Fig. 4 and see text), acts on four different visual pigments, peaking at 530, 550, 570 and 590nm (continuous curves). The absorption by the visual pigments (dotted curves) is obtained by multiplying the absorption spectrum of rhodopsin by the filter transmittance spectrum. (B) Normalizing the resulting spectra yields the sensitivity spectra. (C) Four different red filters (dashed curves) act on a visual pigment peaking at 570nm (continuous curve), giving rise to absorption spectra of lower magnitude (dotted curves). (D) Normalization yields the sensitivity spectra. reflectance spectrum by assuming that the absorption of the visual pigment can be neglected and that the reflectance spectrum of the tapetum is flat in the wavelength range 550–650nm. The normalized reflectance in that range then approximates a modified hyperbolic curve: R(λ)= 1/[1+(λh/λ)n]; where λ is the wavelength of the light. The wavelength of half-reflectance, λh, and the exponent, n, are obtained by fitting the experimental data, yielding values of 590nm and 60, respectively. The normalized transmittance is then calculated by taking the square root of R(λ). The visual pigment in R9 is unknown, so four different rhodopsins are considered, peaking at 530, 550, 570 and 590 nm. Because the rhabdom length of R9 is short, the sensitivity spectrum in the unfiltered situation is virtually identical to the (normalized) absorption spectrum of the rhodopsin, whose shape can be assumed (Stavenga et al., 2000b). Multiplying the filter transmittance spectrum by the rhodopsin absorption spectrum and subsequent normalization yields the sensitivity spectrum of R9 (Fig. 5B). The induced spectral shift depends on the rhodopsin spectrum. The sensitivity peak wavelengths are bathochromic-shifted relative to the rhodopsin peak wavelengths by 44, 37, 27 and 16 nm, respectively. Fig. 5A shows that the resulting sensitivity depends strongly on the overlap between the rhodopsin and filter spectra. Of course, the absolute sensitivity is enhanced by the reflecting tapetum, but this will never amount to more than a factor 2. Fig. 5C depicts a photoreceptor with a rhodopsin peaking at 570nm and the spectral shifts induced by four red filters with λh values of 590, 610, 630 and 650nm (n=60). The induced spectral shifts are 27, 38, 46 and 51nm, respectively. It is again clear that rhodopsin and filter spectra should have at least some overlap, as both the increment in spectral shift and the absolute sensitivity progressively drop when the overlap severely declines. 1.0 A 1.0 C 0.5 0.5 00 1.0 B 1.0 D 0.5 0.5 00 500 550 600 650 700 500 550 600 Wavelength (nm) 650 700 The small white Pieris rapae has two types of red-filtering pigment (Qiu et al., 2002), with λh values of approximately 610 and 640nm. The induced spectral shifts in the red- and deep-red-pigmented ommatidia will be distinctly different. As indicated by Fig. 5, the shifts will depend strongly on the visual pigments in the corresponding R9 photoreceptors. We would expect P . rapae to have rhodopsins peaking near 600 nm because the rhodopsin and filter spectra will then overlap. Of course, the screening pigments will also induce spectral shifts in the proximal photoreceptors, R5–R8, but these shifts may be less pronounced because the pigment is distributed along an extended part of the rhabdom. As shown for Papilio xuthus, a more detailed analysis of the spectral effects of the screening pigments will be greatly facilitated when the sensitivity spectra of the various photoreceptor types are known (Arikawa et al., 1999a,b); such an analysis will then also allow the function of the tapetal mirrors to be assessed. Function of red sensitivity Shifting the sensitivity spectrum of a photoreceptor with a short-wavelength-absorbing filter is well known, e.g. the oil droplets in bird cones (Govardovskii, 1983) and the carotenoid filters in stomatopod rhabdoms (Marshall et al., 1991; for a review, see Douglas and Marshall, 1999). Filtering inevitably causes a reduction in absolute sensitivity, but this cost can be reduced by the tapetal mirror, and it can be easily worth the benefit of enhanced colour contrast discrimination (Govardovskii, 1983). The red receptors of butterflies may be of special importance during oviposition for discriminating suitable leaves for the larvae (Bernard and Remington, 1991; Chittka, 1996; Kelber, 1999). The extremely dense red pigmentation in the Pieridae and the apparently dual system for enhancing red sensitivity strongly suggest that spectral discrimination in the red part of the spectrum is especially Sensitivity AbsorbancePDF Image | Reflections on colourful ommatidia of butterfly eyes
PDF Search Title:
Reflections on colourful ommatidia of butterfly eyesOriginal File Name Searched:
butterfly-eyes.pdfDIY PDF Search: Google It | Yahoo | Bing
Cruise Ship Reviews | Luxury Resort | Jet | Yacht | and Travel Tech More Info
Cruising Review Topics and Articles More Info
Software based on Filemaker for the travel industry More Info
The Burgenstock Resort: Reviews on CruisingReview website... More Info
Resort Reviews: World Class resorts... More Info
The Riffelalp Resort: Reviews on CruisingReview website... More Info
| CONTACT TEL: 608-238-6001 Email: greg@cruisingreview.com | RSS | AMP |